Rivista di formazione e aggiornamento di pediatri e medici operanti sul territorio e in ospedale. Fondata nel 1982, in collaborazione con l'Associazione Culturale Pediatri.
Novembre 2011 - Volume XIV - numero 9
M&B Pagine Elettroniche
Il punto su
L'autismo e il sistema dei neuroni specchio
Ex
falso sequitur quodlibet
DICA
- Laboratorio di Bioingegneria Industriale, Dipartimento di
Ingegneria Civile e Architettura, Università di Udine
Indirizzo
per corrispondenza:bioing@uniud.it
|
Autism
and the mirror neuron system
Key
words
Autism
spectrum disorders, Motor programming deficit evidence, Mirror
neuron, Broken mirror theory
Summary
The
large general consent gained by mirror
neuron paradigm has led to advocate this neural network in the
physiopathology of autism spectrum disorders (ASD). This
hypothesis, tough, is based on a controversial neurophysiologic
assumption: an inappropriate attribution of an emergent cognitive
function to a specific neural population. 'Mirror neuron'
could be the name given to the impossibility to reduce
action-perception dualism to the unity of consciousness.
|
Vari e
vasti sono i campi d'indagine esplorati nel tentativo di
raggiungere un'uniforme e condivisa interpretazione della
fisiopatologia e dei meccanismi che sottendono i disturbi dello
spettro dell'autismo (autism spectrum disorders, ASD),
ovviamente con l'obiettivo, un giorno, di poter disporre di mezzi
efficaci per il trattamento.
Tra i
differenti approcci impiegati per raggiungere un simile risultato,
l'indagine psicologica ambisce a distinguere la sorgente della
distorsione percettiva che conduce al complesso sintomatico. Una
volta distinti differenti sintomi e segni, ricondotti alle diverse
aree dello sviluppo coinvolte,
si è assegnata a essi una giustificazione in termini cognitivi
giungendo a identificare l'assenza di empatia1-4,
ossia la capacità di stabilire un contatto emotivo con gli altri,
come la principale responsabile degli atteggiamenti che
contraddistinguono i disturbi dello spettro autistico. Dai primi
resoconti di un deficit dell'abilità imitativa evidenziata da
Rogers e Pennington nei primi anni '905,6
si giunse a formulare l'ipotesi di una carenza, o isolamento,
emozionale in questi individui tale da determinare l'incapacità di
attuare un processo mentale di immedesimazione, astrazione necessaria
per poter consentire un atto cognitivo complesso come quello
dell'emulazione cosciente. In sostanza, stando a questa
interpretazione, i bambini autistici non sarebbero in grado di
imitare in quanto recepiscono come alieno chi dovrebbero imitare, non
essendo in grado di stabilire con questo un contatto emotivo.
A tal proposito va precisato che l'empatia stessa potrebbe essere
attenuata in conseguenza dell'incompiuta e deficitaria formazione
della 'teoria della mente' nei soggetti autistici (TOM)7-9.
Con tale concetto si designa la capacità più ampia di un individuo
di costruirsi immagini meta-rappresentazionali dei processi cognitivi
altrui partendo dagli stessi propri, ossia individuare nell'altro
in una determinata condizione lo stesso ordine e funzionamento dei
modelli cognitivi ai quali il soggetto stesso ricorre se posto nella
medesima situazione.
La
rappresentazione 'embodied' delle azioni tratteggiata dalla
teoria del sistema dei neuroni specchio (Mirror Neuron System,
MNS)10 richiama da vicino un simile meccanismo psicologico
che consente, nell'uniformità dei processi cognitivi,
l'immedesimazione con gli altri (ossia l'empatia) attraverso la
mappatura diretta tra l'azione osservata e quella eseguita dal
soggetto stesso (direct matching hypothesis), a patto che
naturalmente sia preservata l'integrità della programmazione
motoria11. Da ciò emerge la proprietà cognitiva di
distinzione del fine di ogni singola azione motoria sin dai primi
atti del suo svolgimento. In tale ottica sia quei sintomi riguardanti
la sfera delle interazioni sociali (la risposta alla mimica facciale,
la prossemica, la fissazione con lo sguardo) sia quelli come il
ritardo ad apprendere il linguaggio o la difficoltà a cogliere il
punto di vista di chi sta parlando, che coinvolgono la sfera della
comunicazione verbale, potrebbero dunque sottendere un simile, se non
lo stesso, difetto cognitivo.
Questa
idea, già paventata da psichiatri e psicologi tempo addietro, ha
guadagnato maggior attenzione e consistenza da quando i neuroni
specchio sono stati descritti. Questi, infatti, per le proprietà a
essi attribuite, dovrebbero rappresentare il trait d'union tra la
dimensione personale e quella interpersonale, consentendo il
riconoscimento dell'altro come realtà non aliena ma simile nelle
forme di percezione-azione: proprio ciò che appare deficitario in
soggetti con ASD. Per la prima volta, dunque, a teorie psicologiche
suffragate solo da esperimenti comportamentali hanno potuto essere
affiancate 'inoppugnabili' ricerche neurofisiologiche propense a
spiegare l'abilità imitativa dell'uomo, così come la formazione
della teoria della mente, l'empatia e il linguaggio.
Pertanto,
l'ammissibilità ancora dibattuta dell'esistenza di un sistema
mirror nel cervello umano12-15
precede l'ipotesi che un non corretto funzionamento di questo
sistema possa essere responsabile degli aspetti più caratteristici
dei disturbi dello spettro dell'autismo. In ultima analisi, dunque,
mirror e autismo costituiscono una congiunzione logica
soggetta al principio 'ex falso sequitur quodlibet'.
Recentemente,
proprio un disordine di organizzazione del MNS (che potrebbe
coinvolgere tanto la materia grigia quanto quella bianca) è stato
sondato da Oberman16 come responsabile in prima battuta
del quadro sintomatologico, tanto da aver coniato per questa
l'immaginifico appellativo di 'broken mirror theory'. Il
nodo su cui verte una simile riflessione è che tra soggetti
caratterizzati da un normale sviluppo cognitivo e bambini autistici
debba esistere una differente capacità nell'organizzazione delle
catene di atti motori. Simile evenienza è stata poi descritta
confrontando l'esecuzione di un'azione coordinata come quella di
afferrare del cibo e portarlo alla bocca in questi due distinti
gruppi17. In breve, l'esperimento consisteva nel
monitorare il gesto di afferrare del cibo (pulsante di avvio e
piattaforma sensibile al tocco, su cui veniva collocato il cibo) e
confrontarlo con le tempistiche di attivazione del muscolo
miloioideo, principale attuatore dell'apertura della bocca
(registrato con metodica elettromiografica). Per il gruppo di bambini
autistici si era osservata un'eccitazione del muscolo in ritardo
rispetto al momento in cui il cibo veniva raggiunto con la mano; la
spiegazione offerta in termini neuro-cognitivi suggeriva che la
disgregazione delle catene cinematiche rendesse conto della
difficoltà di distinguere il fine, ossia l'ultimo atto delle
catene motorie, nei soggetti affetti da ASD.
Tuttavia,
gli stessi esperimenti riproposti hanno fornito risultati
diametralmente opposti18, suggerendo l'evenienza che
l'approccio sperimentale impiegato non abbia tenuto conto di una
serie di aspetti critici messi in evidenza
in Figura
1.

Si
osservi innanzitutto come il grafico riporti i risultati dei due
gruppi di soggetti autistici all'interno del range di normalità
demarcato dai risultati dei due gruppi di individui con normale
sviluppo cognitivo.
Per
interpretare una simile prova, bisogna prendere atto che le azioni
motorie sono selezionate e affinate secondo criteri di economia
energetica caratterizzate da precise variabili cinematiche19.
Si può intuitivamente ritenere che l'istante di apertura della
bocca sia necessariamente legato alla rapidità di esecuzione
dell'azione motoria del sistema mano-braccio. In tal senso,
vertendo l'ottimizzazione del movimento principalmente su
coordinate spazio-temporali, sarebbe lecito supporre che
l'attivazione del muscolo mieloioideo, nel momento in cui si
adempie un compito motorio simile a quello descritto
nell'esperimento, debba necessariamente anticipare la propria
azione attraverso un modello predittivo che gestisca il flusso di
informazioni somatosensitive, intrecciando gli schemi dei due atti
motori compresi nell'azione. Simile considerazione obbliga
necessariamente a rapportare l'esecuzione del gesto a quelle che
sono le sue variabili cinematiche per assegnare valore ai distinti
pattern riscontrati all'elettromiografia. Questa considerazione
rende immediatamente conto della discordanza tra quanto descritto da
Cattaneo e coll.17 e quanto da noi rilevato alla luce
della sottostima nel modello sperimentale dei parametri cinetici.
A riprova
di ciò, eseguendo un esperimento analogo su soggetti sani in cui il
target sia collocato di volta in volta a distanze inferiori o
superiori allo spazio esplorabile dal sistema mano-braccio è
identificabile la stretta dipendenza che intercorre tra questo
parametro e la tempistica di apertura della bocca18.

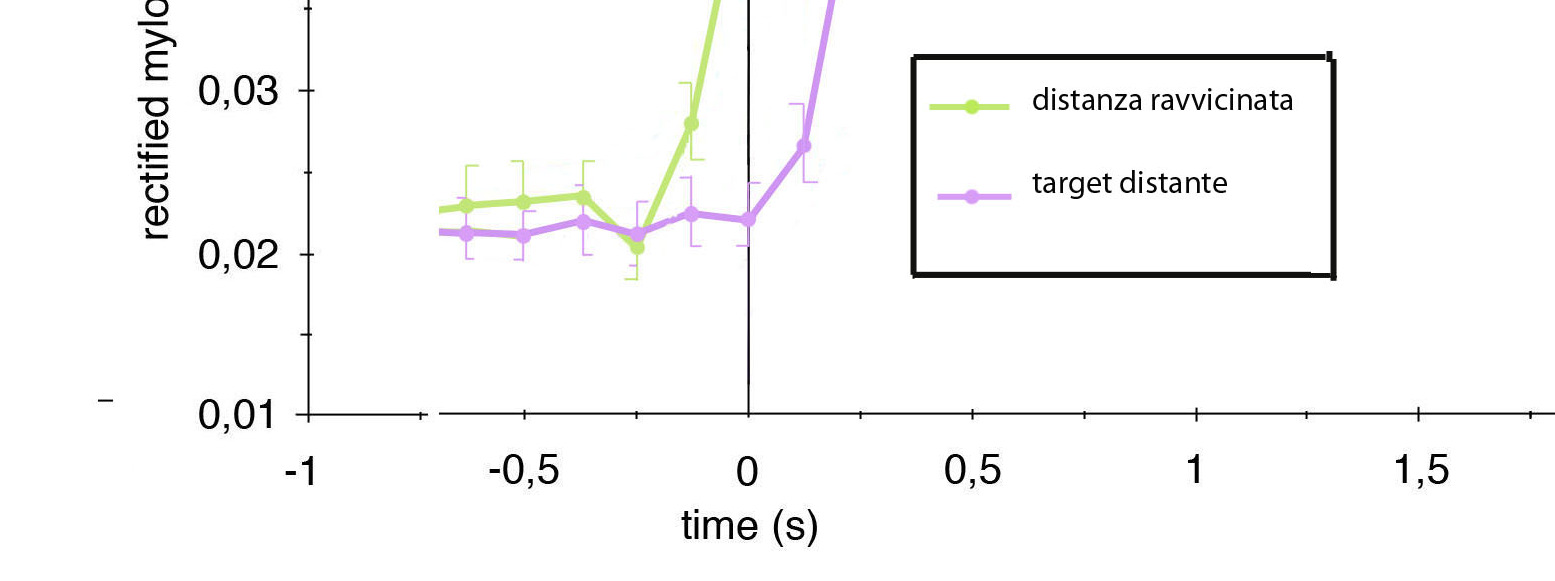
Figura
2. Le due curve rappresentano la media dei risultati di 60 prove
eseguite su un gruppo di bambini con normale sviluppo cognitivo. Il
'bersaglio' viene collocato di volta in volta a una distanza
normalizzata per la lunghezza del braccio del soggetto inferiore o
superiore a quella dello spazio esplorabile dal sistema mano-braccio.
Si noti che l'apertura della bocca avviene in ritardo (curva viola,
a destra) quando il cibo viene collocato a una distanza aumentata.
L'istante 0 indica il momento in cui la mano del soggetto raggiunge
il cibo.
Dall'analisi
delle curve che rappresentano la dinamica di apertura della
bocca (Figura
2), considerato
come istante 0 il raggiungimento dell'obiettivo (criterio di
sincronizzazione di tutte le prove), appare evidente che il movimento
di apertura della bocca si abbia con un maggiore ritardo, nell'ordine
dei 150 ms quando il target viene presentato a una distanza
aumentata. Ciò porta immediatamente a decretare come impropria
l'assunzione che l'attivazione del muscolo miloioideo possa
rappresentare l'espressione di una cognizione del fine dell'azione
motoria innescata, su cui tutte le riflessioni di Cattaneo e coll.
implicitamente si basano. Non si può certamente ritenere che il
'goal', inteso come conclusione dell'azione motoria innescata,
possa in alcun modo dipendere dalla distanza dall'obiettivo. Se
tale assunzione fosse valida, il grafico precedente dimostrerebbe che
in un soggetto sano lo scopo dell'azione già avviata si
costituirebbe in fieri e la sua comparsa parrebbe indiscutibilmente
legata alla distanza del soggetto dall'obiettivo. Conclusione,
questa, insostenibile.
Risulta
altreì più ragionevole ritenere che un simile parametro venga
assunto per adattare la strategia motoria subordinata al
raggiungimento dello scopo già prefigurato. L'apertura della
bocca, conformemente a quanto ipotizzato e come dimostrato in
quest'ultima prova, è diretta espressione delle caratteristiche
cinematiche del gesto, e un parametro come la distanza dal target
(ampiamente trascurato nei precedenti esperimenti descritti), che
direttamente condiziona le variabili associate all'esecuzione del
gesto (impulso, accelerazione massima e velocità massima in primis),
risulta essere decisivo e centrale nel determinare il tempismo con
cui si attiva la muscolatura (genioioideo, miloioideo e digastrico)
che presiede all'apertura della bocca.
In
conclusione, alla luce di quanto emerso dalle rilevazioni da noi
eseguite, si può affermare con certezza che un modello sperimentale
come quello a cui sino ad ora si è ricorsi per sondare l'efficienza
della programmazione motoria nei bambini autistici, come causa della
sintomatologia centrale di questa sindrome, presenta profonde lacune
metodologiche che necessariamente discreditano le conclusioni basate
su risultati sperimentali qui apertamente messi in discussione. Se da
un lato emerge chiaramente che le caratteristiche cinematiche,
grandezze fisiche opportunamente quantificabili, dell'esecuzione
del gesto non sono state sino ad ora appropriatamente tenute in
considerazione nel disegno delle prove sperimentali esaminate,
dall'altro appare ragionevole e auspicabile che a una rigorosa
valutazione di queste si debba ricorrere qualora questa ipotesi, per
molti versi suggestiva, voglia essere seriamente investigata. A tal
proposito appare evidente che un deficit in una simile abilità di
organizzazione di catene motorie può essere riscontrato proprio
valutando la plasticità di adattamento degli schemi motori al
modificarsi di quelle variabili su cui il sistema effettore deve
basarsi per determinare e organizzare traiettorie efficaci.
In ultima
analisi, appare dunque necessaria la strutturazione di nuove batterie
di test in cui la versatilità di pianificazione e l'abilità di
esecuzione di movimenti finalizzati 'goal oriented' vengano messe
alla prova ricorrendo a condizionamenti ambientali esterni posti ad
uopo dallo sperimentatore.
Si
ritiene inoltre auspicabile un comune criterio di reclutamento dei
candidati a questi test, in modo che sia poi possibile una
comparazione delle evidenze su scala più ampia. I limiti
comunicativi che tali soggetti riscontrano, o, vista dall'altro
lato, che noi riscontriamo nel rapportarci a loro, rendono tuttavia
remota la possibilità che anche individui colpiti da sintomatologia
più severa e maggiormente caratteristica possano prendervi parte.
Come
chiaramente emerso dalla nostra esperienza, inoltre, enorme cura
andrà posta nel ridurre al minimo l'impatto delle condizioni al
contorno dell'esperimento, visto il peso non trascurabile che esse
rivestono nel determinare il risultato finale anche su soggetti sani.
Qualora poi un tale deficit venisse riscontrato, l'attribuzione di
questo a una particolare area o popolazione neuronale, in virtù
della complessità e integrazione organizzativa del sistema motorio,
e delle molteplici anomalie morfo-strutturali identificate in aree
concorrenti alla pianificazione-esecuzione motoria in soggetti ASD da
più Autori20-25, appare evidente essere una forzatura,
ancor più considerando quali e quanti siano i dubbi che avvolgono il
modello dei neuroni specchio26.
In
conclusione, l'interessamento del MNS nelle sindromi dello spettro
autistico rimane tuttora da dimostrare. In tal modo si ritiene di
aver fugato la possibile distorsione logica operata da quanti a
riprova dell'esistenza di un simile network nell'uomo ricorrano
all'esemplificazione dell'autismo come dimostrazione di un
mancato funzionamento dello stesso.
1.
Baron-Cohen S, Leslie AM, Frith U. Does the
autistic child have a 'theory of mind'? Cognition
1985;21:37-46.
2.
Baron-Cohen S, Ring HA, Wheelwright S, et al. Social intelligence in
the normal and autistic brain: an fMRI study. Eur J Neurosci
1999;11:1891-8.
3.
Baron-Cohen S, Jolliffe T, Mortimore C, Robertson M. Another
advanced test of theory of mind: evidence from very high functioning
adults with autism or asperger syndrome. J
Child Psychol Psychiatry 2002;38:813-22.
4.
Baron-Cohen S, Golan O, Ashwin E. Can
emotion recognition be taught to children with autism spectrum
conditions? Philos Trans R Soc Lond B Biol
Sci 2009;364:3567-74.
5.
McEvoy RE, Rogers SJ, Pennington BF.
Executive function and social communication deficits in young
autistic children. J Child Psychol Psychiatry 1993;34:563-78.
6.
Rogers SJ, Bennetto L, McEvoy R, Pennington BF. Imitation
and pantomime in high-functioning
adolescents with autism spectrum disorders.
Child Dev 1996;67:2060-73.
7.
Lind SE, Bowler DM. Language and theory of
mind in autism spectrum disorder: the relationship between complement
syntax and false belief task performance. J
Autism Dev Disord 2009;39:929-37.
8.
Lind SE, Bowler DM. Delayed
self-recognition in children with autism spectrum disorder.
J Autism Dev Disord 2009;39:643-50.
9.
Brne M, Brne-Cohrs U. Theory of
mind-evolution, ontogeny, brain mechanisms and psychopathology.
Neurosci Biobehav Rev 2006;30:437-55.
10.
Gallese V. Intentional
attunement: a neurophysiological perspective on social cognition and
its disruption in autism.
Brain Res 2006;1079:15-24
11.
Iacoboni M, Woods RP, Brass M, Bekkering. H, Mazziotta JC, Rizzolatti
G. Cortical mechanisms of human imitation. Science 1999;286:2526-8.
12.
Hickok G. Eight problems for the mirror
neuron theory of action understanding in monkeys and humans. J
Cogn Neurosci 2009;21:1229-43.
13.
Dinstein I, Hasson U, Rubin N, Heeger DJ. Brain
areas selective for both observed and executed movements.
J Neurophysiol 2007;98:1415-27.
14.
Pascolo PB, Budai R. Neuroni mirror nell'area F5 della corteccia
cerebrale della scimmia: c'è stata evidenza sperimentale? E
nell'uomo? New Magazine Edizioni. Rivista
Medica. 2008;60.
15.
Pascolo PB, Budai R, Rossi R. Critical review of the research leading
to the mirror neuron paradigm - biomed 2010. Biomed Sci Instrum
2010;46:422-7.
16.
Oberman LM, Ramachandran VS. The simulating
social mind: the role of the mirror neuron system and simulation in
the social and communicative deficits
of autism spectrum disorders. Psychol
Bull 2007;133:310-27.
17.
Cattaneo L, Fabbri-Destro M, Boria S, et al. Impairment
of actions chains in autism and its possible role in intention
understanding. Proc Natl Acad Sci USA 2007;104:17825-30.
18.
Pascolo PB, Ragogna P, Cremaschi S, et al. Autism
and motor acts: experimental analysis on mylohyoid muscle emg
recordingsduring grasping-to-eat action - biomed 2010. Biomed Sci
Instrum 2010;46:178-83.
19.
Nelson WL. Physical principles for economies of skilled movements.
Biol Cybern 1983;46:135-47.
20.
Courchesne E, Carper R, Akshoomoff N. Evidence of brain overgrowth in
the first year of life in autism. JAMA 2003;290:337-44.
21.
Courchesne E, Karns CM, Davis HR, et al. Unusual brain growth
patterns in early life in patients with autistic disorder: an MRI
study. Neurology 2001;57:245-54.
22.
Hashimoto T, Tayama M, Murakawa K, et al. Development of the
brainstem and cerebellum in autistic patients. J
Autism Dev Disord 1995;25:1-18.
23.
Keller TA, Kana RK, Just MA. A
developmental study of the structural integrity of white matter in
autism. Neuroreport 2007;18:23-7.
24.
Waiter GD, Williams JH, Murray AD, Gilchrist A, Perrett DI, Whiten A.
Structural white matter deficits in high-functioning individuals with
autistic spectrum disorder: a voxel-based investigation. Neuroimage
2005;24:455-61.
25.
Zilbovicius M, Garreau B, Samson Y, et al. Delayed maturation of the
frontal cortex in childhood autism. Am J Psychiatry
1995;152:248-52.
26. Fan
YT, Decety J, Yang CY, Liu JL, Cheng Y. Unbroken mirror neurons in
autism spectrum disorders. J Child Psychol Psychiatry 2010;51:981-8.
Vuoi citare questo contributo?